
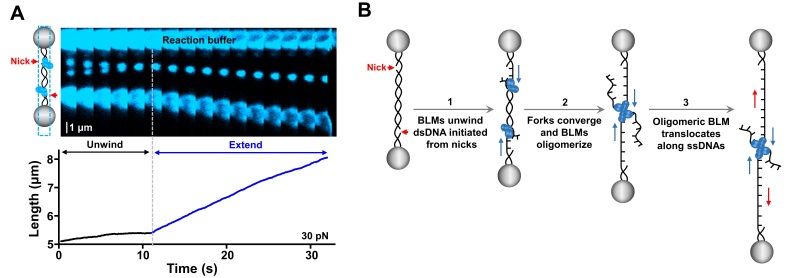
图1:A, BLM对向解旋并聚集后实现单链DNA滑动;B, BLM解旋酶活性转换模型。
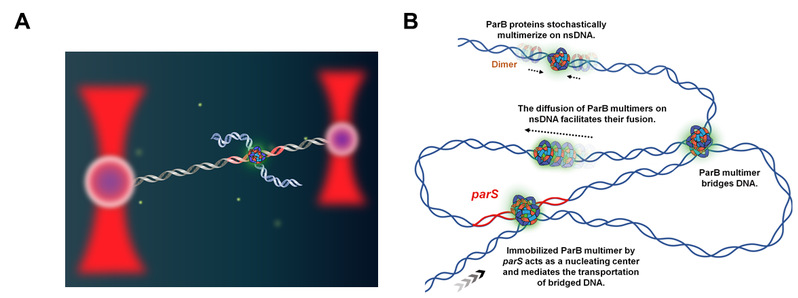
图2:单分子实验及ParB多聚体组织DNA组装的分子机制模型。
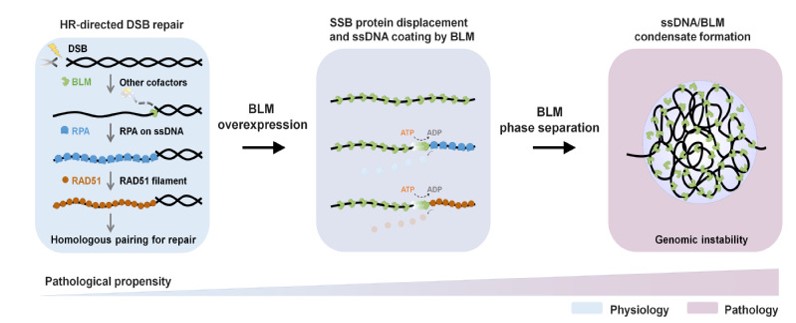
图3:BLM过表达影响同源重组,导致基因组不稳定模型。
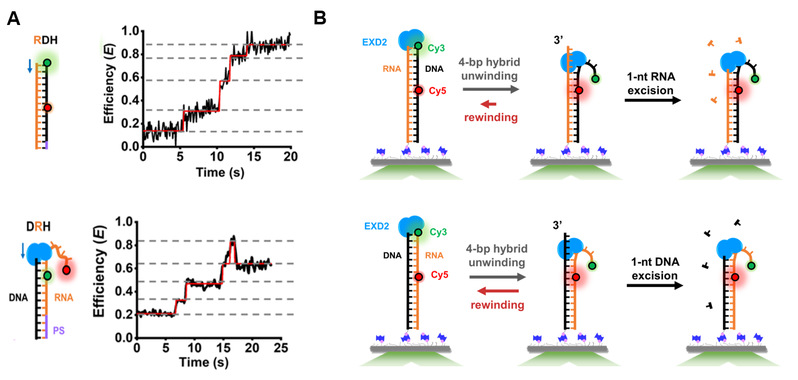
图4:EXD2切割RNA–DNA杂合体的动力学分析及模型。
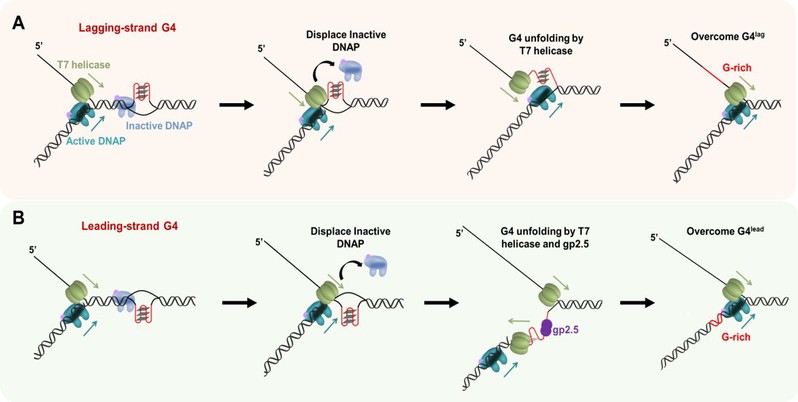
图5:T7复制体克服前导链和后随链G-四链体的分子机制。
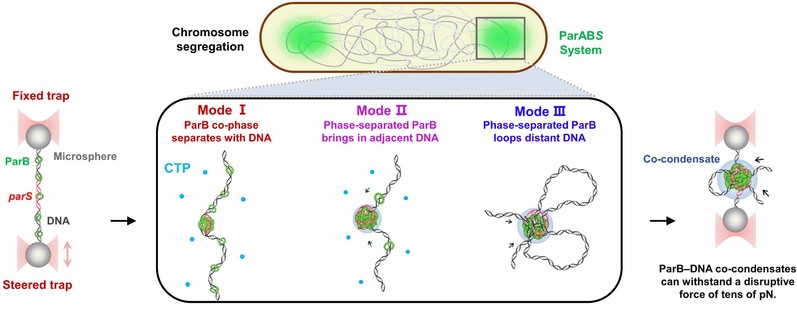
图6: 染色体分离蛋白ParB通过相分离组装染色体分离复合物。
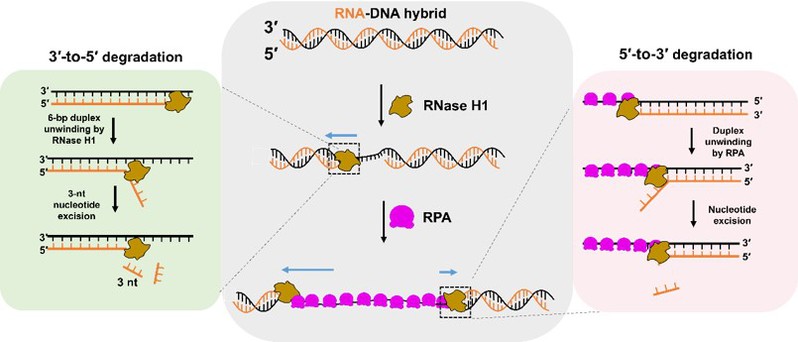
图7:RNase H1催化双向RNA-DNA杂合核酸降解的分子机制模型。
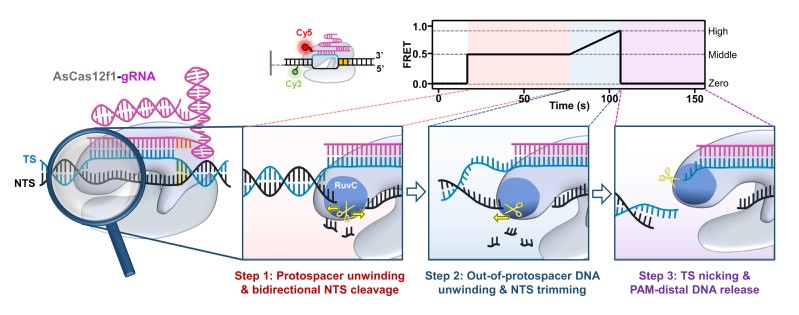
图8:AsCas12f1催化DNA靶序列顺序切割的分子机制模型。
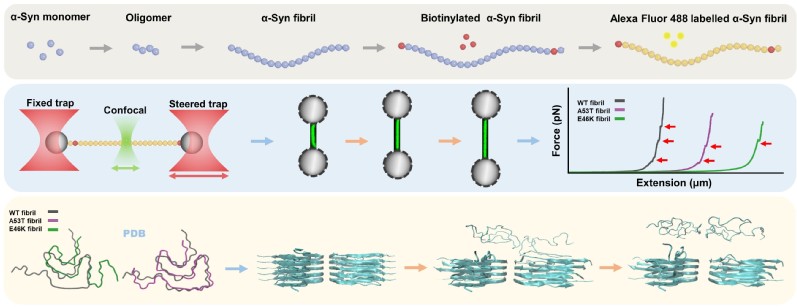
图9:Alpha-syn淀粉样纤维的单分子检测及分子动力学模拟。
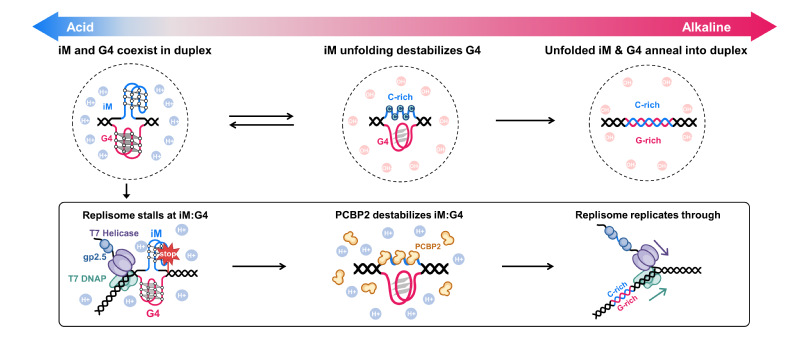
图10:iM:G4结构的动态变化及PCBP2介导的复制耐受机制。
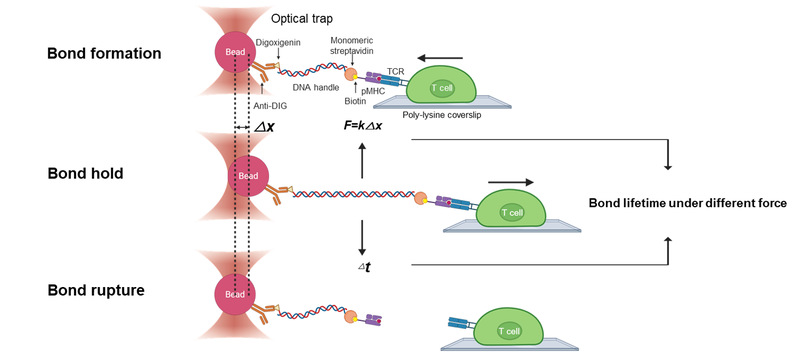
图11:单分子光镊技术检测TCR-pMHC逆锁键强度。
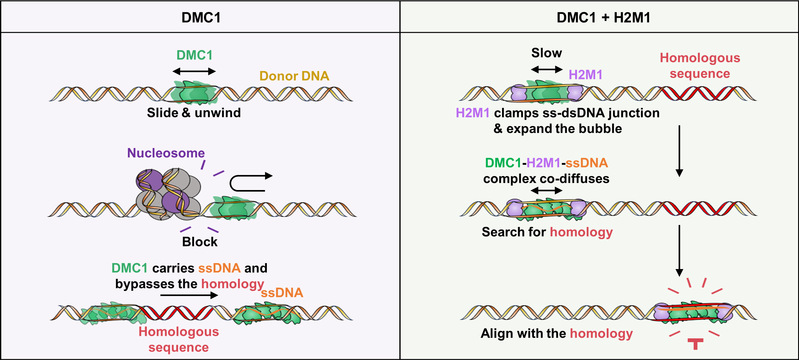
图12:重组酶DMC1在H2M1的帮助下精准识别同源序列模型。
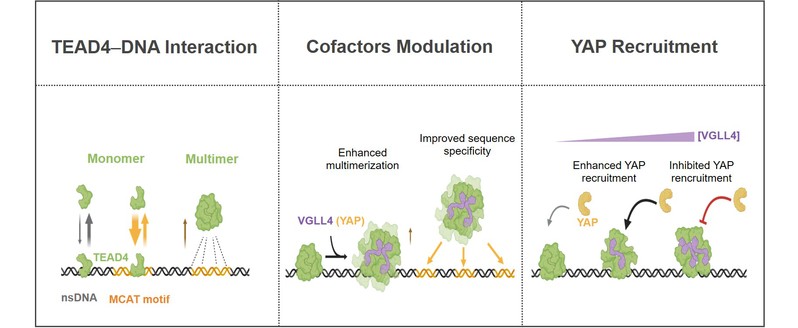
图13: VGLL4调控Hippo转录因子TEAD4结合DNA及招募YAP模型。