
Recently, CRISPR-derived base editors have been developed to realize precise base conversion in DNA. Cytosine base editor (CBE) can induce base conversion from cytosine (C) to thymine (T) at on-target sites. However, early-developed CBEs can also trigger gRNA-dependent DNA off-target (OT) mutations and gRNA-independent DNA OT mutations, which hinder their therapeutic applications.
In order to comprehensively eliminate the OT mutations induced by CBEs, Prof. Jia Chen's research team at ShanghaiTech University and their collaborators have developed a highly precise CBE, named transformer base editor (tBE), which triggers no observable gRNA-dependent DNA OT mutations, gRNA-independent genome-wide OT mutations or gRNA-independent transcriptome-wide OT mutations.
Based on this research, Prof. Jia Chen's research team and Prof. Jianfeng Li's research team at ShanghaiTech University, and Prof. Li Yang's research team at the Center for Molecular Medicine, Children’s Hospital, Fudan University jointly published a protocol paper in Nature Protocols, entitled Design and application of the transformer base editor in mammalian cells and mice. The paper describes the working principle, design scheme, and in vivo editing by the tBE.
The paper first describes how tBE works in detail (Figure 1). The tBE system consists of four proteins, including (1) a Streptococcus pyogenes Cas9 (SpCas9) nickase D10A (nSpCas9), (2) the N-terminal fragment of tobacco etch virus (TEV) protease (TEVn), (3) a fusion protein of an RNA aptamer binding protein (N22p) and the C-terminal fragment of TEV protease (TEVc) and (4) an effector fusion protein called tBE–V5–mA3 that contains a cytidine deaminase (mouse apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3 (APOBEC3) cytidine deaminase domain 1, mA3CDA1), a deoxycytidine deaminase inhibitor (dCDI) from mouse APOBEC3 (mA3dCDI), a uracil DNA glycosylase inhibitor (UGI) and another RNA aptamer binding protein (MCP) (Fig. 1a). In addition, the tBE system also includes a boxB aptamer-containing single-guide RNA (sgRNA–boxB) and an MS2 aptamer-containing helper sgRNA (hsgRNA–MS2) (Fig. 1a).
When the tBE system is expressed in the cytoplasm of cells, its cytidine deaminase is inactive due to the linkage between mA3dCDI and mA3CDA1 via the TEV protease cleavage site (Fig. 1b, cytoplasm), which ensures that no gRNA-dependent or gRNA-independent OT mutations are triggered. When nSpCas9-hsgRNA and nSpCas9-sgRNA are binding to the on-target site simultaneously, hsgRNA-MS2 recruits the effector protein tBE-V5-mA3, whereas sgRNA-boxB recruits the fusion protein of N22p-TEVc (Fig. 1b, colocalization). A free TEVn and TEVc recruited by sgRNA-boxB combine to form an intact TEV protease, which cleaves off the mA3dCDI that is fused to cytidine deaminase via the TEV protease cleaving site (Fig. 1b, activation). Therefore, mA3CDA1’s cytidine deaminase activity is activated at the on-target site and able to induce efficient base editing (Fig. 1b, base editing). Because a split version of the TEV protease is used in the tBE system, the trimolecular reaction leading to cytidine deaminase activation of mA3dCDI occurs only at the on-target site (Fig. 1b). At any OT site (gRNA-dependent or gRNA-independent), random collisions of any two molecules of TEVn, TEVc with the TEV protease cleavage site do not result in cytidine deaminase activation, thus preventing the generation of OT mutations. Moreover, only two active cytidine deaminases are theoretically released from the two alleles of a target site following the successful on-target editing by tBE as SpCas9 dissociates slowly from the on-target site.
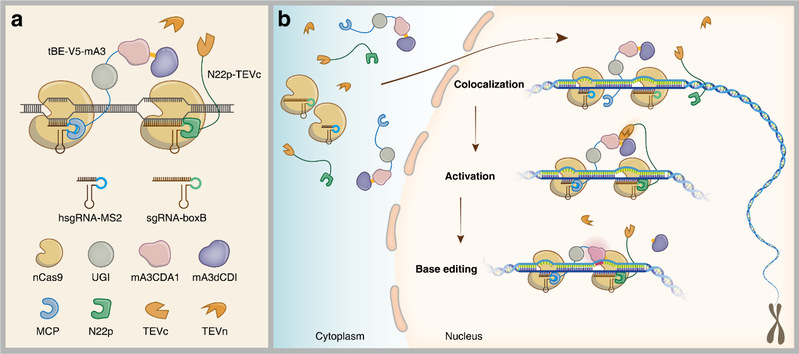
Figure 1. Components (1a) and mechanism (1b) of tBE.
The paper also introduces the design of sgRNA-hsgRNA pair. The position selection of the sgRNA-hsgRNA pair is very critical when utilizing tBE for gene editing, generally, when the hsgRNA located in the 34-91 bp region upstream of the sgRNA, this sgRNA-hsgRNA pair combination can produce optimal editing efficiency at the on-target site. The hsgRNAs should be designed to target the strand that the sgRNA targets, which can avoid the generation of insertions or deletions (indels). After screening for the optimal sgRNA-hsgRNA pair for the on-target site in commonly used cell lines, the authors provide a detailed description of the experimental method of utilizing a dual adeno-associated virus to package the tBE system and deliver it to mice to detect the therapeutic effect.
Next, the article also provides comprehensive details on how to detect gRNA-dependent DNA OT mutations, gRNA-independent genome-wide OT mutations and gRNA-independent transcriptome-wide OT mutations, lists experimental methods and data analysis tools.
In summary, compared with other CBE systems, tBE is a well-designed and accurate base editor without observable OT events, which provides a safe and efficient tool for precision gene editing therapy.
In this paper, Prof. Jia Chen, Prof. Jianfeng Li from ShanghaiTech University and Prof. Li Yang from the Center for Molecular Medicine, Children’s Hospital, Fudan University are the co-corresponding authors. Dr. Wenyan Han from Prof. Jia Chen's lab, Dr. Baoqing Gao from Prof. Li Yang's lab, Dr. Junjie Zhu from Prof. Jia Chen's lab, and Dr. Zongxing He from Prof. Jianfeng Li are the co-first authors.
Paper link: https://www.nature.com/articles/s41596-023-00877-w